Защитное действие нейролипинов при гипоксии
Влияние нейролипинов на жизнеспособность нейронов в условиях гипоксии изучали на культурах нейронов новой коры и гиппокампа головного мозга крысы. Нейроны коры помещали на 4 часа в камеру, продуваемую смесью азот/углекислота (95/5%), после чего инкубировали 16 часов в нормальной атмосфере. В данных условиях наблюдалось увеличение гибели нейронов в 1,5-1,6 раза по сравнению с культурами, находившимися в условиях нормоксии, о чем свидетельствовало соответствующее повышение активности лактатдегидрогеназы (ЛДГ) в среде инкубации. В данных условиях все исследованные нейролипины анандамид, 2АГ, АД, ДД) проявили защитное действие (рис. 12). Важно отметить, что эффект наблюдался при более низких концентрациях соединений (0,01-1 мкМ) по сравнению с другими моделями.

Воздействие гипоксии на жизнеспособность и функциональные свойства нейронов также изучали на клетках, выделенных из гиппокампа крысы и культивируемых на мультиэлектродных матрицах. Выживаемость определяли подсчетом живых и погибших клеток при помощи микроскопии после соответствующего окрашивания. Функциональную активность нейронов определяли регистрацией внеклеточных потенциалов на электродах мультиэлектродной матрицы.
Влияние нейролипинов и их рецепторов на указанные параметры оценивали на примере АДА. Было установлено, что через сутки после короткой гипоксии (10 мин в атмосфере аргона) во всех исследованных группах наблюдалось снижение жизнеспособности нейронов в среднем на 15%. На седьмые сутки после гипоксии жизнеспособность нейронов в контрольной группе снизилась до 30,8%, тогда как на фоне АДА (10 мкМ) количество живых клеток не изменилось по сравнению с первыми сутками после гипоксии. Антагонисты KP1 и BP1 снижали защитное действие АДА (табл. 4).

АДА (10 мкМ); SR1 — антагонист KP1 SR141716A (2,5 мкМ); SR2 — антагонист КР2 SR144528 (2,5 мкМ); Кап — антагонист BP1 капсазепин (2,5 мкМ); * р<0,05 сравнение с нормоксией, ** р<0,05 сравнение с 1-ми сутками после гипоксии тест Стьюдента
В культурах нейронов гиппокампа, подвергнутых гипоксии, также наблюдалось изменение характера спонтанной сетевой активности. В контрольных культурах происходило увеличение среднего количества сетевых пачек через 10 минут после гипоксии (рис. 13А). Количество внеклеточных потенциалов (ВП) в пачке оставалось при этом неизменным (рис. 13Б). Количество пачек снижалось через 2 часа и через сутки после гипоксии, а на 7-е сутки наблюдалось практически полное прекращение пачечной активности. Также отмечалось снижение количества ВП в пачке после гипоксии на первые и седьмые сутки. Аппликация АДА (10 мкМ) препятствовала развитию изменений, наблюдаемых в контроле, и приводила к сохранению структуры пачечной активности сети в первые сутки постгипоксического периода (см. рис. 13А). Важно отметить, что использование АДА приводило к сохранению жизнеспособности клеток на 7-е сутки на уровне первых суток после гипоксии (см. табл. 4). Таким образом, наблюдаемое снижение биоэлектрической активности на фоне АДА связано с функциональными изменениями, а не со снижением количества живых клеток, как в контрольных культурах.

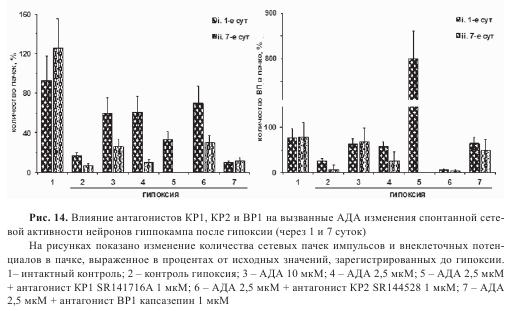
Для выявления рецепторного компонента в реализации эффектов АДА были использованы антагонисты KP1-, КР2- и BP1-рецепторов. Блокада KP1-рецепторов на фоне действия АДА приводила к снижению количества пачек через сутки после гипоксии (рис. 14А). Также наблюдалось выраженное увеличение (примерно в 8 раз) количества ВП в пачке, что свидетельствует о растормаживании сети за счет блокады KP1 рецепторов (рис. 14Б). На 7-е сутки после гипоксии при блокаде KP1 происходило полное исчезновение сетевой активности. При блокаде BP1 на 1-е и 7-е сутки количество пачек уменьшалось, тогда как число ВП в пачке оставалось на том же уровне, что и у АДА. При блокаде КР2-рецепторов не происходило достоверных изменений количества пачек на 1-е и 7-е сутки по сравнению с действием АДА, при этом количество ВП в пачке резко снижалось на 1-е сутки и оставалось далее неизменным (см. рис. 14А,Б).
Важно отметить, что блокада KP1- и BP1-рецепторов снижает защитное действие АДА на 7-е сутки (см. табл. 4). Таким образом изменение биоэлектрической активности на 7-е сутки в присутствии антагонистов KP1 и BP1 также может быть связано с уменьшением количества жизнеспособных нейронов. Полученные данные свидетельствуют о том, что АДА обладает выраженным нейропротекторным действием в условиях гипоксии, а также способствует сохранению функционального состояния нейронов гиппокампа в постгипоксический период. На регуляцию сетевой активности в постгипоксический период наиболее заметное влияние оказывают KP1-рецепторы. Тем не менее, общее влияние АДА на биоэлектрическую активность и выживаемость культивируемых клеток гиппокампа, по-видимому, является результатом тонкой настройки сетевой активности в результате суммарного взаимодействия со всеми тремя типами рецепторов.
Читайте также
Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа

Когда начинать готовиться к ЕГЭ и ОГЭ 2024: полезные рекомендации

Начало нового учебного года часто становится временем повышенной тревожности как
На чем можно долететь до Мальдив? Регулярный рейс или аренда частного самолета?

Путешествие на Мальдивы — это мечта многих туристов. Острова, утопающие
Зубной имплантат: преимущества выбора при протезировании

Зубной имплантат – это современная технология, предоставляющая возможность восстановить утраченный
Яйцо шоколадное Kinder сюрприз: волшебство, которое завоевало сердца детей и взрослых

Яйцо Kinder сюрприз, безусловно, является одним из наиболее популярных шоколадных
Суши и пицца: почему они так популярны в службе доставки

Службы доставки еды становятся всё популярнее среди людей, желающих насладиться
Пептидные препараты: сущность и области применения

Пептидные препараты стали одним из важнейших направлений в современной медицине