Влияние нейролипинов на нейрональный апоптоз
Модель калиевой депривации является одним из распространенных способов изучения нейронального апоптоза. Нейроны мозжечка крысы сохраняют жизнеспособность при культивировании в среде с повышенным содержанием ионов калия (25 мМ). При понижении концентрации калия до 5-10 мМ в течение 18-24 часов наблюдается массовая гибель нейронов по пути апоптоза. В данной модели были протестированы аналоги анандамида, 2АГ, ACT и АДА. В исходной постановке опыта все вещества исследовали в трех концентрациях: 0.1, 1,10 мкМ. Анандамид, 2АГ и ACT, а также их аналоги в данном тесте оказались неактивны. Среди N-ацилдофаминов нейрозащитное действие проявили АДА и N-докозагексаеноилдофамин (ДДА, 1а-5e). Максимальный эффект наблюдали при концентрации 10 мкМ: 59% и 39% выживших клеток для АДА и ДДА, соответственно. Для этих соединений было проведено исследование зависимости защитного эффекта от концентрации (рис. 4).
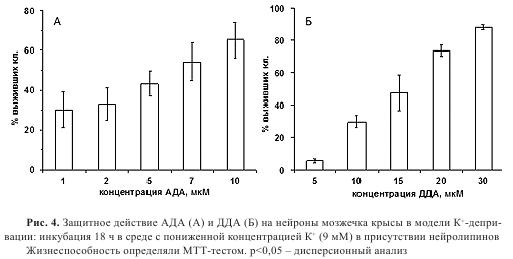
АДА увеличивал выживаемость нейронов в диапазоне концентраций 1-15 мкМ (30-65% выживших клеток), тогда как ДДА был активен в интервале 5-30 мкМ (10-85% выживших клеток). Применение антагонистов каннабиноидных, ванилоидных и дофаминовых рецепторов показало, что эффекты N-ацилдофаминов реализуются преимущественно за счет КР2 и BP1 (рис. 5). Аналоги АДА с модификациями в дофаминовой части не оказывали достоверного защитного действия.
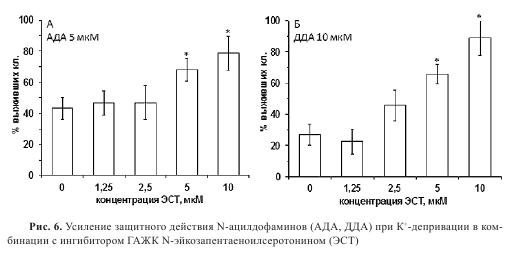
С учетом биохимических характеристик нейролипинов, описанных выше, нами была применена комбинация соединений, состоящая из нейролипина и ингибитора его гидролиза. Были использованы АДА и ДДА в комбинации с наиболее активным ингибитором ГАЖК N-эйкозапентаеноилсеротонином (ЭСТ, 1с-4b).

Было установлено, что ЭСТ снижает эффективную концентрацию АДА и ДДА в условиях К+-депривации. В присутствии АДА (5 мкМ) и ДДА (10 мкМ) выживание клеток повышалось на 30-40% по сравнению с контролем. На фоне повышающейся концентрации ЭСТ 1-10 мкМ наблюдали усиление нейропротекторного эффекта АДА и ДДА до 80-90% (рис. 6). Эффект комбинации ДДА+ЭСТ заметно уменьшался в присутствии антагониста КР2, тогда как антагонист KPl на него не влиял.
Характерными признаками развития апоптоза являются активация кальций-зависимых протеаз (каспаз) и фрагментация ядерной ДНК.
Электрофоретический анализ ДНК, выделенной из клеток, показал, что при инкубации с АДА и ДДА количество фрагментов с характерной длиной 180 пар оснований заметно снижается по сравнению с контролем (данные не приведены). Оценку активности каспаз 3 и 9 в культуре нейронов проводили с использованием флуоресцентно меченых субстратов. Было показано, что при К+-депривации увеличивается активность обеих каспаз, что свидетельствует в пользу развития гибели нейронов по пути апоптоза в данной модели. В присутствии АДА и ДДА активность этих ферментов снижена в 4-5 раз (рис. 7), что служит подтверждением антиапоптотического действия этих соединений.

Для выяснения возможных путей внутриклеточной сигнализации, задействованных в защитном эффекте АДА и ДДА, проведен ингибиторный анализ ряда клеточных протеинкиназ. Для этого АДА и ДДА применяли в сочетании с ингибиторами фосфоинозитол-3-киназы (PI3K), протеинкиназы A (PkA), протеинкиназы С (PkC), калций-кальмодулин-зависимой киназы 2-го типа (CaMK-II), киназ, активируемых внеклеточными сигналами (МЕКК, MEK-1,2). В присутствии ингибиторов PI3K, PkC и CaMK-II наблюдали выраженное снижение нейропротекторного действия АДА и ДДА, что может свидетельствовать в пользу участия этих протеинкиназ во внутриклеточных каскадах, приводящих к повышению жизнеспособности нейронов под действием этих нейролипинов (рис. 8).

Читайте также
Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа

Когда начинать готовиться к ЕГЭ и ОГЭ 2024: полезные рекомендации

Начало нового учебного года часто становится временем повышенной тревожности как
На чем можно долететь до Мальдив? Регулярный рейс или аренда частного самолета?

Путешествие на Мальдивы — это мечта многих туристов. Острова, утопающие
Зубной имплантат: преимущества выбора при протезировании

Зубной имплантат – это современная технология, предоставляющая возможность восстановить утраченный
Яйцо шоколадное Kinder сюрприз: волшебство, которое завоевало сердца детей и взрослых

Яйцо Kinder сюрприз, безусловно, является одним из наиболее популярных шоколадных
Суши и пицца: почему они так популярны в службе доставки

Службы доставки еды становятся всё популярнее среди людей, желающих насладиться
Пептидные препараты: сущность и области применения

Пептидные препараты стали одним из важнейших направлений в современной медицине