Структурная организация моторного контроля
Основными звеньями системы регуляции моторного поведения являются сенсомоторная кора, мозжечок, красное ядро и система стриатума. Каждая из этих структур имеет свою специфику в анализе периферической афферентации и формировании моторной команды, которая реализуется через кортикоспинальную и другие нисходящие системы мозга. В определенной степени к звеньям системы регуляции моторного поведения также можно отнести четверохолмие, дающее начало тектоспинальной системе и формирующее, в частности, старт-рефлекс на внезапные зрительные и слуховые раздражения, и ретикулоспинальную систему, являющуюся одним из основных непирамидных нисходящих выходов.
Согласно классическим представлениям, план движения возникает в ассоциативных областях коры и реализуется через мозжечок, формирующий программу быстрых движений, и базальные ганглии (для медленных движений) и моторную кору. Существенная роль отводится кортико-мозжечковым петлям. При этом считалось, что моторная кора контролирует спинальные мотонейроны по принципу «точка в точку», как система «высших мотонейронов».
Современные исследования существенно изменили эти представления. Показано, что активность нейронов моторной коры коррелирует с разными параметрами движения, включая скорость, силу, направление движения в суставах, конечное положение конечности. Микростимуляция моторной коры может вызывать сложные движения. Предполагается, что моторная система использует метод «оптимального контроля» множества параметров при выполнении каждого конкретного движения. Показано участие моторной коры в контроле позы и в обучении новой позе.
Существенную роль играет пластичность и изменение функций разных отделов моторной коры и других структур в процессе обучения и при компенсации мозговых повреждений. Показано, что при обучении в моторной коре происходит активный синаптогенез.
Мозжечок играет существенную роль в моторном программировании. Латеральный мозжечок получает информацию о плане движения от ассоциативных областей коры и формирует программу движения, которая через зубчатые ядра и ядра таламуса передается на премоторную и моторную кору, где формируется нисходящая моторная программа, копия которой через нижнюю оливу поступает в промежуточную и медиальную области мозжечка, где она сравнивается с афферентной информацией. Корректирующий сигнал передается на спинной мозг через соответствующие ядра мозжечка, красное ядро и вестибулярные ядра. Через последние мозжечок участвует в контроле позы и равновесия. Показана также роль мозжечка в позной подготовке к движению и в организации ритмических движений.
Роль мозжечка в обучении была теоретически обоснована Г.С. Бриндли, Д. Марром и Дж.С. Олбусом и затем показана экспериментально в ряде исследований. Обнаружен синаптогенез в мозжечке при обучении. Показано участие мозжечка в реорганизации двигательных координаций в процессе обучения.
Считается, что мозжечок формирует внутреннюю модель тела и объектов окружающей среды, которая используется в процессе обучения. Согласно «вычислительной гипотезе обучения», существует три вида обучения — обучение «с учителем», т.е. по обратной связи (supervised learning), обучение по подкреплению (reinforcement learning) и обучение по ассоциации (unsupervised learning). Мозжечок играет существенную роль в обучении по обратной связи, оценивая текущую ошибку движения, тогда как функция базальных ганглиев связана, в основном, с выбором реакции для достижения максимума подкрепления (reinforcement learning). Моторная кора, согласно этой гипотезе, вовлечена в обучение по ассоциации. Показано, однако, что мозжечок, базальные ганглии и моторная кора вовлечены «в обучение с учителем» (supervised learning) в ситуации обучения произвольному контролю положения центра тяжести по зрительной обратной связи.
Базальные ганглии, и в частности дофаминергическая система среднего мозга, привлекают наибольшее внимание в аспекте настоящей работы. Согласно современным представлениям, важную роль в функциях базальных ганглиев играет, во-первых, дофаминергический вход из компактной части черной субстации, нейроны которой, как и дофаминергические нейроны вентральной тегментальной области, реагируют на награду в ситуации обучения, и, во-вторых, сложная организация выхода из стриатума (прямой и непрямой пути), обеспечивающая выбор реакции («принятие решения») и торможение ненужных движений. Стриатум получает также билатеральный кортикальный вход через нейроны переднего мозга и ипсилатеральный вход через коллатерали пирамидных нейронов. Показано, что конвергенция кортикального и дофаминового входов в дорзальном стриатуме облегчает организацию поведения, направленного на получение подкрепления, причем D1- и D2-рецепторы на нейронах стриатума могут играть разную роль в зависимости от позитивного или негативного подкрепления. Через прямой выход (D1-рецепторы) стриатум тормозит внутренний паллидум и ретикулярную часть черной субстанции, уменьшая их тормозящие влияния на таламо-кортикальные входы (т.е. растормаживает их). Таким образом, прямой выход является активирующим. С другой стороны, через непрямой выход (D2-рецепторы) стриатум тормозит внутренний паллидум и снижает его тормозящее влияние на субталамическое ядро, что приводит к увеличению активирующего влияния последнего на внутренний паллидум и ретикулярную часть черной субстанции и в результате увеличивает их тормозящие влияния на таламо-кортикальные входы и уменьшает растормаживающее влияние прямого пути. Таким образом, непрямой выход является тормозным (рис. 1).
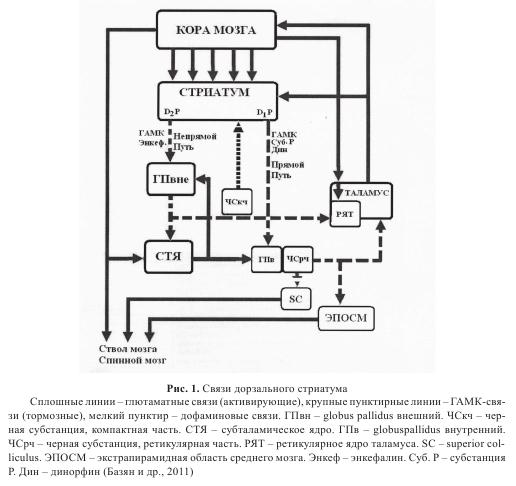
Базальные ганглии являются частью экстрапирамидной системы мозга, участвующей в моторном контроле. Центральной структурой базальных ганглиев является стриатум, состоящий из трех ядер. Прилежащее ядро, или nucleusaccumbens, относят к вентральному стриатуму. Nucleus caudatus, или хвостатое ядро, и putamen, или скорлупа, относятся к структурам дорзального стриатума и составляют неостриатум.
95% всех нейронов дорзального стриатума — это ГАМКергические средние звездчатые нейроны. Остальные 5% — это интернейроны: большие незвездчатые нейроны составляют 1-2% клеточной популяции стриатума. Они используют ацетилхолин как медиатор. Третий тип нейронов — это средние незвездчатые нейроны. Это также интернейроны, они используют соматостатин как передатчик. Четвертый тип нейронов — это маленькие незвездчатые нейроны, которые используют ГАМК как медиатор. Всего интернейронов 5%, что очень мало для функционирования локальной сети. Нехватка интернейронов в дорзальном стриатуме компенсируется специальным строением аксонов и дендритов средних звездчатых нейронов стриатума, что позволяет им участвовать в функционировании локальной сети. Одиночный аксон средних звездчатых нейронов дорзального стриатума, идущий к нейронам паллидума и ретикулярной части черной субстанции, обладает коллатералями у самого основания аксона; они иннервируют дендритное дерево собственного и соседних средних звездчатых нейронов. Следовательно, средние звездчатые нейроны сами участвуют в локальной обработке информации дорзальным стриатумом, используя ГАМК как медиатор.
Иерархические сети дорзального стриатума имеют специфическое строение. Стриатум получает возбудительный вход практически от всех областей коры головного мозга (см. рис. 1). Корковый вход использует глутамат как собственный медиатор и заканчивается в значительной степени на головках дендридных шипиков средних звездчатых нейронов. Проекции от коры головного мозга к стриатуму имеют топографическую организацию. Например, соматосенсорная и моторная кора проецируются к заднему неостриатуму, а префронтальная кора проецируется к переднему хвостатому ядру. Внутри соматосенсорных и моторных проекций к стриатуму сохраняется соматотопия. Было предположено, что топографические отношения между корой головного мозга и стриатумом обеспечивают базу для изоляции функционально различных сетей в базальных ганглиях (рис. 2). Эти циркулирующие сети включают соматомоторные, глазодвигательные и лимбические связи. Внутри каждой сети предполагается наличие подсети, причем таким образом, чтобы у первичной моторной коры и премоторной коры были неидентичные связи со структурами базальных ганглий. Аналогично, дорзолатеральные и орбитолатеральные сети обладают особыми соединительными паттернами.

Из рис. 2 видно, что активность из коры приходит к определенным частям хвостатого ядра или неостриатума, оттуда — к определенным частям внутреннего сегмента globuspallidus (или эндопендикулярного ядра у грызунов) и ретикулярной части черной субстанции, оттуда — к определенным ядрам таламуса и обратно в соответствующую область коры. Мы полагаем, что наличие параллельных разделенных сетей позволяет также понять механизм стереотипного поведения, который может функционировать наподобие параллельных сетей. Например, стереотипия круговых движений, челночных движения, жевания, лизания, грызения и тому подобное. К такому роду стереотипии можно отнести и реакцию самостимуляции, которая возникает в определенных условиях, при активации префронтальной коры. В эти поведенческие стереотипии вовлекается и nucleusaccumbens — ядро вентрального стриатума.
Хотя топография и соматотопия подразумевают определенную степень параллельной организации, есть также конвергенция и дивергенция в кортико-стриатных проекциях. Большие дендритные области средних звездчатых нейронов позволяют им получать вход от смежных проекций, которые приходят из различных областей коры. Входы больше чем от одной корковой области перекрываются, и вход от одиночной корковой области проецируется дивергентно к многочисленным зонам стриатума, например, сенсорно-двигательные корковые проекции — к стриатуму от областей руки в соматосенсорной коре (области 1, 2, и 3) и моторной коры (область 4). Таким образом, каждая корковая область проецируется к нескольким стриатальным зонам, и несколько функционально связанных корковых областей проецируются к одной стриатальной зоне. Эта сходящаяся и расходящаяся организация служит анатомической основой для интеграции и преобразования информации от нескольких областей коры головного мозга, в том числе и при условнорефлекторном обучении. Эти дивергентые и конвергентные связи могут интегрировать разные представительства в коре мозга, в том числе и представительства условного и безусловного раздражителей.
Показано, что холинергическая система стриатума имеет отношение к контролю позного компонента выученного движения. Эти исследования являются развитием классических представлений о роли базальных ганглиев в контроле позы. Показана связь дофаминергических и холинергических механизмов стриатума в его интегративной деятельности. При этом обнаружены независимые дофаминергические системы управления позой и движением. Существенная роль принадлежит также взаимодействию холинергической и серотонинергической систем стриатума. Показаны различия нарушений обучения при поражениях стриатума и мозжечка: мозжечок преимущественно участвует в подготовке к движению, а нарушения функций нигро-стриатной системы при болезни Паркинсона приводят к нарушениям внутренней оценки и выбора.
Обнаружена пластичность кортико-стриатных входов. Синаптическая пластичность регулируется дофамином. После денервации стриатума путем ипсилатеральной корковой ишемии происходит мощный рост аксонов контралатеральных кортико-стриатных нейронов. В начальной стадии паркинсонизма возникает дефицит стриато-фронтальных проекций, что приводит к активации связей дорзальной премоторной коры с моторной корой.
Согласно современным представлениям, принятие решения и выбор правильной реакции для получения подкрепления обусловлены функциональной активностью дофаминергической системы. Показано, что уровень дофамина в нейронах среднего мозга изменяется в соответствии с вероятностью стимула предшествовать подкреплению. Фазические ответы дофаминергических нейронов возникают при неожиданном подкреплении. После повторных сочетаний условного стимула с подкреплением эти ответы начинают возникать при предъявлении условного стимула. Активность дофаминергических нейронов тормозится при неподкреплении условного стимула. Таким образом, дофаминергическая система кодирует возможную ошибку получения подкрепления и способствует обучению поведению, приводящему к максимуму подкрепления. Это поведение хорошо соответствует вычислительной модели. Через рецепторы D1, D2, D3, D4 и D5 дофамин снижает активность непрямого выхода из стриатума и повышает активность прямого выхода.
Читайте также
Особенности сантехники из Италии: почему стоит переплатить

Отдых в Джемете — солнце, море, пляж

Джемете — это живописный район на побережье Черного моря, который
Школа профессионального макияжа: искусство создания идеального образа

-

Дорамы – популярный жанр телевизионного и кинематографического искусства, который нашел
Правильный выбор ткани — половина успеха вашего образа

Разработка брендбука: почему это важно

Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа
