Применение блокаторов ионотропных рецепторов при пентилентеразоловом киндлинге
Целью настоящей части работы было исследование морфологических изменений в нейронах гиппокампа мышей, возникающих в результате киндлинга, и возможной корреляции между способностью блокаторов глутаматных рецепторов NMDA-типа мемантина, ИЭМ-1921 и ИЭМ-1958, предупреждать как судорожные проявления киндлинга, так и сопутствующие морфологические изменения. Была использована модель ПТЗ киндлинга, т.е. состояния повышенной судорожной готовности, возникающей в ходе систематических (с интервалом в 2-3 дня) введений подпороговой дозы ПТЗ, блокатора ГАМК-ергической синаптической передачи.
Исследование возможной корреляции между судорожными проявлениями ПТЗ киндлинга и структурными изменениями нейронов гиппокампа было проведено на пяти группах мышей: (1) интактные мыши; (2) контрольные мыши, которым с целью развития киндлинга по стандартной схеме вводили ПТЗ; (3) мыши, которым вводились 0,3 мг/кг ИЭМ 1921 + ПТЗ; (4) мыши, которым вводились 1 мг/кг ИЭМ-1958 + ПТЗ и (5) мыши, которым вводились 0,5 мг/кг мемантина + ПТЗ.
Первое введение ПТЗ мышам контрольной группы не вызывало судорожных реакций. Лишь слабые подергивания кончиков ушей и вибрисс были отмечены у части животных. Начиная со второго введения, у единичных мышей (10-20% животных) возникали слабо выраженные и непродолжительные клонические судороги. По мере увеличения порядкового номера последовательных введений такой же дозы ПТЗ процент мышей, проявивших судорожные припадки, нарастал, достигая максимума (70%) к 11—12-ому введению (рис. 6). К концу эксперимента у 70% мышей контрольной группы наблюдались клонико-тонические судороги (ср. балл 3,9), а 30% мышей оказались устойчивыми к действию ПТЗ и не судорожили на протяжении всего опыта (ср. балл 0). Общий средний балл у мышей контрольной группы ПТЗ киндлинга составил 2,7.
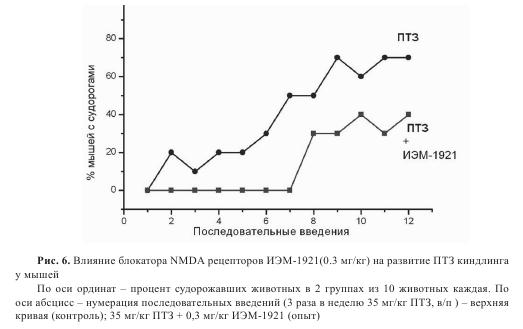
У экспериментальной группы мышей, которым на протяжении месяца за 30 мин до стандартной подпороговой дозы ПТЗ вводили 0,3 мг/кг ИЭМ-1921, развитие киндлинга существенно замедлилось. У 14 из 20 мышей, взятых в опыт, судорожная реакция полностью отсутствовала после первых 7 введений ПТЗ и постепенно появлялась, начиная с 8-ого введения, устойчивый ПТЗ-киндлинг был достигнут лишь к концу эксперимента (12 введений ПТЗ + ИЭМ-1921). У 6 остальных мышей судорожные проявления отсутствовали до конца эксперимента. Средний балл интенсивности судорог в группе (ПТЗ+ИЭМ-1921) составил 0,7, т.е. в 3,9 раза ниже, чем в контрольной группе.
Используя аналогичную схему, было проведено исследование другого нового блокатора глутаматных рецепторов NMDA типа, ИЭМ-1958, производного адамантана. У 5 из 8 мышей этой серии ПТЗ киндлинг не развился до конца эксперимента. У остальных 3 мышей первые судорожные припадки силой 2-3 балла появились после 9-ого введения, т.е. профилактическое действие препарата оказалось весьма эффективным. Мемантин был использован в настоящем исследовании как препарат сравнения. При его введении в дозе 0,5 мг/кг у 5 мышей из 8 ПТЗ киндлинг не развился.

В контрольной группе животных киндлинг вызывал существенные структурные изменения пирамидных нейронов поля СА1 гиппокампа. Если у интактных животных в срезах гиппокампа было обнаружено только 0,15% гиперхромных нейронов (рис. 7а,б), то в контрольной группе мышей с выраженными проявлениями ПТЗ киндлинга (присутствие клонико-тонических судорог) были обнаружены выраженные патологические изменения (рис. 7в). В слое stratum pyramidale поля СА1 гиппокампа содержалось большое количество гиперхромных нейронов (24,7±2,1%). У них наблюдались следующие изменения — потеря тургора, сморщивание и уменьшение размеров клеток, искривление дендритных отростков. Часть клеток принимала удлиненно-суженную форму. При исследовании подгруппы мышей, у которых несмотря на систематическое введение ПТЗ не развился типичный киндлинг, гиперхромные нейроны в срезах гиппокампа были единичны (2,3±2,3%) (см. рис. 7).
Морфологические различия между животными с выраженным киндлингом и устойчивыми к киндлингу были найдены и в экспериментальных группах мышей, которым вводили блокаторы NMDA рецепторов. В мозге животных, устойчивых к киндлингу, не было обнаружено каких-либо патологических изменений нейронов, тогда как у подверженных киндлингу животных гиперхромные нейроны присутствовали. При сравнении нейропротекторного действия различных препаратов у животных с выраженными судорогами было выявлено, что ПЭМ-1921 слабо предотвращал появление гиперхромных нейронов (24,0±5,6%), тогда как введение ПЭМ-1958 и мемантина существенно уменьшало их число (6,6±0,9% и 1,7%, соответственно).
Киндлинг является удобным экспериментальным приемом, поскольку постепенное нарастание патологического процесса дает возможность проследить последовательность и взаимосвязь его проявлений. Это позволяет использовать ПТЗ киндлинг в качестве адекватной модели как при оценке активности противосудорожных средств, так и для выявления дегенеративных изменений нейронов мозга, вызванных судорогами.
Совокупность полученных нами результатов свидетельствовала, что гиперхромные нейроны («темные клетки») появлялись в результате судорожных приступов, перенесенных животными в ходе развития ПТЗ киндлинга. Подобные изменения нейронов «по темному типу» обнаруживались при морфологическом анализе самых разнообразных патологических процессов в мозгу при ишемии, гипогликемии, эпилепсии, избыточном действии возбуждающих медиаторов. Однако существующие мнения о природе этого феномена и даже о его причинно-следственной связи с патологией конкретных нейронов неоднозначны. Часть исследователей даже расценивает появление темных клеток как артефакт, вызванный нарушением условий содержания и фиксации ткани. Тем не менее, проведение надлежащих контролей и получение надежных в каждом отдельном случае признаков связи между гиперхромностью нейронов и патологическим воздействием позволяет использовать этот критерий при оценке интенсивности процесса.
Следующим убедительным аргументом в пользу такой связи можно считать способность трех блокаторов глутаматных рецепторов NMDA типа мемантина, ИЭМ-1921 и ИЭМ-1958 предупреждать развитие не только судорожного компонента ПТЗ-киндлинга, но и появление гиперхромных нейронов в гиппокампе. Молекулярный механизм взаимодействия использованных блокаторов с активируемыми глутаматом NMDA рецепторами подробно исследован. Производные адамантана (мемантии и ИЭМ-1958) и фенилциклогексила (ИЭМ-1921) блокируют потенциалозависимым образом открытое состояние канала NMDA рецептора, конкурируя при этом с ионами магния. Блокаторы отличает высокая степень селективности по отношению к открытым каналам глутаматных рецепторов NMDA типа. Эти особенности молекулярного механизма блокады делают их хорошими инструментами исследования вовлеченности NMDA рецепторов в различные физиологические и патологические процессы.
Ранее нами было проведено сравнение противосудорожной активности мемантина и ИЭМ-1921 с использованием нескольких моделей в опытах на мышах: острые судорожные состояния, вызываемые внутрижелудочковым введением NMDA или каината, внутрибрюшинным введением ПТЗ, ареколина, ПТЗ киндилинг и в опытах на крысах линии Крушинского-Молодкиной, генетически предрасположенных к аудиогенным судорогам. В настоящем исследовании возможной корреляции между противосудорожным и нейропротекторным действием неконкурентных блокаторов NMDA рецепторов были использованы их равноэффективные дозы в диапазоне ЭД40-ЭД55.
Результаты проведенных экспериментов выявили эту корреляцию. В контрольной группе систематическое введение ПТЗ вызывало у 70% мышей состояние киндлинга, что выражалось появлением судорожных ответов. При морфологическом исследовании срезов СА1 гиппокампа этих мышей обнаружено значительное число темных нейронов. Превентивное введение блокаторов NMDA рецепторов снижало долю мышей с развившимся ПТЗ киндлингом в среднем с 70% до 40%. Содержание измененных нейронов у 60% мышей экспериментальных групп, у которых ПТЗ киндлинг не развился и судороги не проявлялись, составило 0,1±0,06%, т.е. столько же, сколько в мозгу интактных мышей. Среди этих 60% могли быть и мыши, резистентные к действию ПТЗ изначально, а не вследствие профилактического действия блокаторов. Однако доля резистентных мышей в контрольной группе была в 2 раза меньше. Кроме того, появление судорожных припадков у опытных групп мышей по сравнению с контрольной было существенно задержано. Даже после 7-го введения ПТЗ ни у одной мыши еще не было судорог, тогда как в контроле они появились уже у 50% животных. Эти результаты позволяют считать, что блокада NMDA рецепторов способна не только замедлять и ослаблять развитие ПТЗ киндлинга, и, соответственно, судорожные реакции, но и предотвращать появление темных нейронов в гиппокампе.
По поводу причинно-следственной связи между судорогами и феноменом темных нейронов существуют противоречивые данные. Прежде всего, появляются ли темные клетки в мозгу крыс после однократного введения дозы 70-80 мг/кг ПТЗ, вызвавшей судороги, либо темные клетки могут быть обнаружены только в результате повторных введений ПТЗ, причем как при исследовании мозга животных с выраженными судорожными проявлениями, так и при их видимом отсутствии. Согласно результатам настоящего исследования ПТЗ киндлинга у мышей, предшествующий судорожный синдром является непременным условием обнаружения темных клеток в гиппокампе. В основе расхождения данных могут лежать различия в объектах (крысы, мыши), оценке интенсивности судорог и критериев морфологических изменений.
Мало известно о дальнейшей судьбе темных клеток после возникновения в мозгу патологического очага, эти сведения необходимы для более полного понимания их природы. В последнее время появляются результаты комплексных исследований процессов дегенерации зрелых нейронов и пролиферации молодых нейронов в ростовых областях нервной ткани. Работа в этом направлении способна пролить свет на общую проблему клеточных механизмов эпилепсии. Требует также уточнения роль глутамата в последовательности процессов, приводящих к повреждениям мозговой ткани, вызванным судорожным синдромом, и, в частности, ограничивается ли она повышением входа ионов кальция в нейроны через открытые ионные каналы глутаматных рецепторов.
Читайте также
Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа

Когда начинать готовиться к ЕГЭ и ОГЭ 2024: полезные рекомендации

Начало нового учебного года часто становится временем повышенной тревожности как
На чем можно долететь до Мальдив? Регулярный рейс или аренда частного самолета?

Путешествие на Мальдивы — это мечта многих туристов. Острова, утопающие
Зубной имплантат: преимущества выбора при протезировании

Зубной имплантат – это современная технология, предоставляющая возможность восстановить утраченный
Яйцо шоколадное Kinder сюрприз: волшебство, которое завоевало сердца детей и взрослых

Яйцо Kinder сюрприз, безусловно, является одним из наиболее популярных шоколадных
Суши и пицца: почему они так популярны в службе доставки

Службы доставки еды становятся всё популярнее среди людей, желающих насладиться
Пептидные препараты: сущность и области применения

Пептидные препараты стали одним из важнейших направлений в современной медицине