Образование флуорофоров — продуктов взаимодействия свободного транс-ретиналя с аминогруппами белка и липидов в фоторецепторных клетках (исследование in vitro на модельных системах)
Избыточное накопление свободного ПТР в фоторецепторной мембране при некоторых патологиях или в случае интенсивного воздействия света приводит к образованию бисретиноидов — продуктов взаимодействия ПТР с аминогруппами фосфатидиэтаноламина и белковых молекул (А2РЕ и др.). Эти бисретиноиды наряду с флуорофорами ЛГ (A2E и др.) также могут вносить вклад в аутофлуоресценцию глазного дна. Изучение спектральных характеристик бисретиноидов фоторецепторных клеток является не менее актуальным, чем изучение флуорофоров ЛГ в РПЭ. Ситуация избыточного накопления свободного ПТР в фоторецепторных мембранах была промоделирована в экспериментах in vitro.
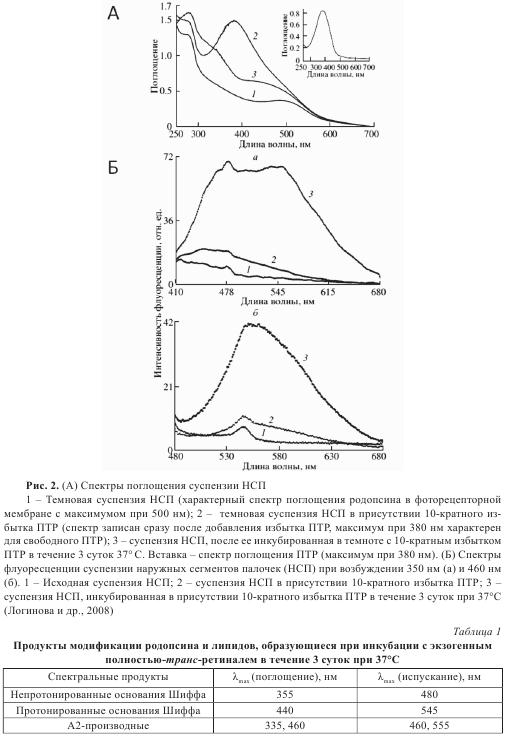
На рис. 2А представлен спектр поглощения (спектр 3) суспензии наружных сегментов палочек сетчатки (НСП) после темновой инкубации с 10-кратным избытком ПТР в течение 3 суток при 37°С. В этот спектр кроме самого родопсина вносят вклад продукты взаимодействия свободного ПТР с аминогруппами белка и липидов.
Продукты взаимодействия ПТР с аминогруппами родопсина и фосфатидилэтаноламина обладают характерной и сильной флуоресценцией. Ниже представлены данные флуоресцентного анализа этих продуктов при длинах волн возбуждения 350 и 460 нм (рис. 2Б). При возбуждении 350 нм наблюдаются две выраженные полосы с максимумами в области 480 нм и 545 нм, а при возбуждении 460 нм — одна полоса с максимумами в области 540-550 нм. Анализ спектров флуоресценции (табл. 1) позволяет предположить, что образуются три типа соединений: согласно литературным данным, это могут быть непротонированные Шиффовы основания, протонированные Шиффовы основания и продукты вторичного взаимодействия ПТР с Шиффовым основанием (например А2-РЕ и А2-родопсин).

Для сравнительного исследования фотохимических свойств транс-ретиналя и его конъюгатов с родопсином и липидами были зарегистрированы кинетики затухания флуоресценции в пикосекундном диапазоне времени после их возбуждения импульсом длительностью ~20 пс с длиной волны 353 нм (рис. 3). Кинетика затухания флуоресценции ПТР в буфере практически моноэкспоненциальна (τ1=31±2 пс) и характеризует время жизни возбужденного состояния. Исследование кинетики затухания флуоресценции суспензии наружных сегментов палочек после инкубации с ПТР в течение 3 суток при 37°С показало, что характер кинетической кривой существенно меняется. Наилучшая аппроксимация этой кинетики достигается в 3-экспоненциальном приближении кривой вида:
Ф(t) = ΣiAi exp(-t/τi),
где i — число компонент, А — амплитуды, τ — длительность компонент. Найдено, что τ1=48±2 пс (22%), τ2=208±5 пс (48%), τ3=900±10 пс (30%). Наличие трех компонент в кинетике флуоресценции инкубированного образца можно объяснить присутствием как минимум 3 центров свечения. Сопоставление с литературными данными позволяет предположить, что первая компонента (48 пс) принадлежит протонированным основаниям Шиффа, вторая компонента (208 пс) принадлежит, по-видимому, А2-производным ретиналя, а третья компонента (900 пс) — непротонированным основаниям Шиффа.
Таким образом, описанная выше модельная система позволяет предположить, что в ситуации избыточного накопления ПТР в фоторецепторной клетке происходит образование как минимум трех типов продуктов. Продукты взаимодействия ПТР с аминогруппами фосфатидилэтаноламина и белковых молекул в фоторецепторных мембранах проявляют достаточно сильные флуоресцентные свойства и могут вносить заметный вклад в аутофлуоресценцию глазного дна, например, при патологиях, вызванных нарушением механизма удаления отработанного ПТР при фотолизе родопсина.
Читайте также
Особенности сантехники из Италии: почему стоит переплатить

Отдых в Джемете — солнце, море, пляж

Джемете — это живописный район на побережье Черного моря, который
Школа профессионального макияжа: искусство создания идеального образа

-

Дорамы – популярный жанр телевизионного и кинематографического искусства, который нашел
Правильный выбор ткани — половина успеха вашего образа

Разработка брендбука: почему это важно

Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа
