Нейролипины — лиганды каннабиноидных рецепторов
Взаимодействие с рецепторами оценивали методом радиолигандного связывания в препаратах плазматических мембран, полученных из клеток и тканей, содержащих соответствующие рецепторы. Для оценки взаимодействия изучаемых нейролипинов с каннабиноидным рецептором 1-го типа (KP1) использовали препараты мембран, выделенные из гомогената головного мозга крысы, а также синтетические радиоактивно меченые лиганды: агонист [3Н]СР55940 и антагонист [3H] SR141716A. На начальном этапе с помощью насыщения рецепторного связывания определяли параметры взаимодействия (Kd) меченых лигандов с рецепторами, а затем проводили оценку связывания нейролипинов и их аналогов методом вытеснения радиоактивно меченых лигандов. Установлено, что наибольшим сродством к KP1 плазматических мембран головного мозга крысы обладают амиды жирных кислот. Анандамид (1d-3а) оказался наиболее эффективным лигандом (Ki=12 пМ) среди эндогенных нейролипинов. Константа ингибирования для N-арахидоноилдофамина (АДА, 1d-5e) составила 100 пМ, тогда как N-арахидоноилсеротонин (ACT, 1d-4b) не проявлял значимой активности (табл. 1).

Замены в жирнокислотной части молекулы АДА на ω3 кислоты с разной длинной алифатической цепи и степенью ненасыщенности приводили к снижению аффинности соответствующих аналогов (см. табл. 1). В семействе дофаминоидов активность соединений в зависимости от структуры жирнокислотного остатка убывала в следующем ряду (приведены обозначения жирных кислот):
C20:4ω6>C22:5ω3>C20:5ω3>C22:6ω3>C18:3ω6»C18:4ω3>C18:3ω3>a-dm-С20:4ω6.
Работами ряда авторов было показано, что замены остатков арахидоновой кислоты в анандамиде и 2АГ на ω3 кислоты приводили к резкому падению аффинности соответствующих аналогов, значения констант ингибирования которых превышали 10 мкМ. Интересно, что у производных дофамина с ω3 жирными кислотами не наблюдается драматических потерь аффинности, тогда как замена арахидоновой кислоты на кислоты С18-семейства независимо от серии (ω3 или ω6) привела к падению аффинности на порядок и более.
Для определения характера взаимодействия с рецептором были проведены исследования специфических для активации KP1 физиологических ответов в тестах на клетках и на животных. Известно, что под действием агонистов KP1 у мышей наблюдается снижение локомоторной активности, каталепсия, снижение ректальной температуры и повышение порога болевой чувствительности. Проявление всех перечисленных ответов под действием исследуемого вещества характеризует его как агонист KP1. Анандамид проявлял выраженную активность во всех тестах тетрады. N-ацилдофамины демонстрировали активность, согласующуюся с результатами экспериментов по связыванию: аналоги, имевшие низкую активность в отношении KP1, также менее эффективны в тетраде (см. табл. 1). Кроме того, было установлено, что N-ацилсеротонины не вызывают характерных для каннабиноидов ответов в проведенных тестах.
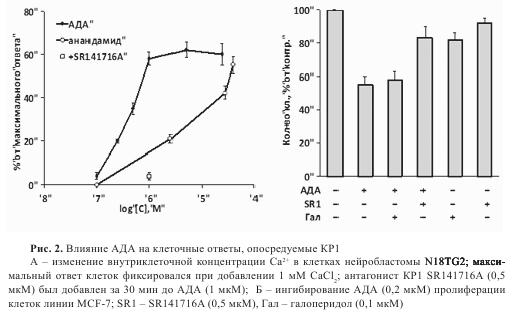
Ранее было показано, что активация KP1, в частности анандамидом, стимулирует в клетках нейробластомы мыши N18TG2 повышение концентрации ионов Ca2+ за счет его выделения из внутриклеточных депо. Нами установлено, что в клетках линии N18TG2 анандамид и АДА стимулируют увеличение концентрации ионов Ca2+ внутри клетки. Причем АДА оказался в 45 раз более активным, чем анандамид (рис. 2). Предынкубация клеток с антагонистом KP1 SR141716A (0,5 мкм, 30 мин) приводила к исчезновению эффекта АДА (1 мкМ). Еще одним характерным ответом на активацию KP1 является ингибирование пролиферации клеток линии рака молочной железы MCF-7. АДА ингибировал пролиферацию с ЕС50=250 нМ, при этом антагонист KP1 SR 141716А препятствовал ингибированию, тогда как неселективный антагонист дофаминовых рецепторов галоперидол не влиял на эффект АДА (см. рис. 2).
Оценку взаимодействия N-ацилдофаминов с каннабиноидными рецепторами 2-го типа (КР2) проводили на препаратах плазматических мембран селезенки крысы, где ранее был обнаружен высокий уровень экспрессии этих рецепторов. В качестве меченого лиганда использовали [3H]WIN-55,212-2. Было показано, что исследуемые соединения имеют низкую аффинность на КР2. Наиболее активным нейролипином из группы амидов жирных кислот оказался анандамид (Ki=3 мкМ). У производных дофамина значения констант ингибирования превышали 10 мкМ.
Ранее было установлено, что некоторые нейролипины (анандамид, 2АГ) способны реализовывать свои эффекты через ванилоидные рецепторы первого типа (ВР1, или TRPV1). АДА в диапазоне концентраций 0,5-25 мкМ не приводил к вытеснению селективного агониста BP1[3H]-резинифератоксина в препаратах мембран спинного мозга крысы.
N-ацилдофамины имеют в своем составе остаток дофамина, что позволяет предположить их взаимодействие с дофаминовыми рецепторами. Однако оказалось, что на мембранных препаратах из мозга крысы АДА (0,1-10 мкМ) не приводил к значимому вытеснению меченых лигандов D1 и D1 рецепторов, [3H]SCH23390 и [3Н]-спиперона, соответственно.
Результаты, полученные в экспериментах по вытеснению меченного лиганда, во многом зависят от его структуры и места его связывания с рецептором, которое может не полностью соответствовать месту взаимодействия на рецепторе исследуемого соединения. Поэтому использование функциональных моделей может дать дополнительную информацию о сигнальных свойствах исследуемых веществ (см. далее). В данной работе, а также другими авторами было установлено, что ряд клеточных ответов, инициируемых АДА, ингибируется антагонистами BP1 и КР2 рецепторов. Таким образом, нельзя исключать, что BP1 и КР2 также являются молекулярными мишенями N-ацилдофаминов.
Полученные результаты на клеточных моделях и в тестах на животных позволяют сделать вывод о том, что N-арахидоноилдофамин и некоторые его гомологи способны реализовывать свои эффекты за счет активации каннабиноидных рецепторов 1-го типа. Важно подчеркнуть, что в составе липидов головного мозга наблюдается повышенное содержание ω3 жирных кислот, преимущественно докозагексаеновой кислоты (С22:6ω3, 1а), которая, как и арахидоновая кислота, может служить предшественником эндогенных нейролипинов. Полученные нами результаты позволяют предположить, что в случае эндогенного образования N-ацилдофаминов, в состав которых входят ω3 кислоты, их молекулярными мишенями также могут быть каннабиноидные рецепторы 1-го типа.
Читайте также
Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа

Когда начинать готовиться к ЕГЭ и ОГЭ 2024: полезные рекомендации

Начало нового учебного года часто становится временем повышенной тревожности как
На чем можно долететь до Мальдив? Регулярный рейс или аренда частного самолета?

Путешествие на Мальдивы — это мечта многих туристов. Острова, утопающие
Зубной имплантат: преимущества выбора при протезировании

Зубной имплантат – это современная технология, предоставляющая возможность восстановить утраченный
Яйцо шоколадное Kinder сюрприз: волшебство, которое завоевало сердца детей и взрослых

Яйцо Kinder сюрприз, безусловно, является одним из наиболее популярных шоколадных
Суши и пицца: почему они так популярны в службе доставки

Службы доставки еды становятся всё популярнее среди людей, желающих насладиться
Пептидные препараты: сущность и области применения

Пептидные препараты стали одним из важнейших направлений в современной медицине