Нейрогенез в мозге при болезни Альцгеймера
Еще до недавнего времени господствовало представление о невозможности нейрогенеза в мозге взрослых млекопитающих и человека, хотя это явление было открыто более 30 лет назад. Сейчас это одна из наиболее «горячих» тем не только нейрофизиологии, но и многих смежных дисциплин. Основными структурами мозга, где происходит нейрогенез во взрослом организме, являются субвентрикулярная зона и зубчатая фасция гиппокампа, а также обонятельная луковица, которую с субвентрикулярной зоной связывает переднемозговой миграционный путь, по которому двигаются нейрональные прогениторы, постепенно дифференцируясь в нейроны, олигодендроциты и астроциты. Показано, что в стареющем мозге наблюдается нарушение нейрогенеза, что может быть одной из причин развития нейродегенеративных заболеваний, включая и БА. Однако при БА обнаружено и усиление пролиферативной активности в мозге. В настоящее время нет полной ясности в этом вопросе ввиду весьма противоречивых данных, полученных в эксперименте на животных моделях БА и в клинике. Исследования с использованием трансгенных животных — моделях БА, не внесли ясности в решение этого вопроса, поскольку в подавляющем большинстве трансгенные животные демонстрировали снижение выраженности нейрогенеза. С другой стороны, модели ускоренного старения у крыс показали усиленный нейрогенез в гранулярных клетках гиппокампа, выраженность которого положительно коррелировала с когнитивным дефицитом. Результаты наших исследований пролиферативной активности в мозге ОБЭ мышей представлены на рис. 7.

В наших исследованиях максимальная пролиферативная активность, которая определялась по включению в клетки мозга BrdU, отмечена в гиппокампе, субвентрикулярной зоне и височной коре ОБЭ животных через 3-4 недели после бульбэктомии, т.е. на фоне развития нейродегенеративных изменений. Действительно, именно в этот период после операции бульбэктомии наблюдали снижение нейрональной плотности у ОБЭ мышей: в височной коре — на 10±1,8%, в поле CA1 гиппокампа — на 18,3±2,6%; в поле САЗ гиппокампа — на 23,8±3,1%, в холинергических структурах переднего мозга (горизонтальном ядре вертикального пучка Брока и гигантоклеточном преоптическом ядре) — на 44,1±9,2%, в хвостатом ядре и скорлупе — на 29±3,1%. Клетки дорзального ядра шва, ответственные за синтез серотонина и обеспечивающие им переднемозговые структуры и кору, резко меняли свою форму и демонстрировали различные формы патологии — пикноз, кариолизис, цитолиз. Число нормальных клеток снижалось до 50,6±10%. Этот факт дает основание думать, что инициатором активации пролиферативной активности в мозге является сама гибель клеток и выделение факторов, компенсаторно усиливающих пролиферативную активность и нейрогенез. Память у ОБЭ животных начинала восстанавливаться только через 12 недель после операции. В это время в гиппокампе и коре были обнаружены нейрональные прогениторы, одновременно иммунопозитивные как к BrdU, так и маркерам разных стадий клеточной дифференцировки, таким как виментин, 3-β-тубулин и NeuN. Наличие клеток с 3-β-тубулином и NeuN указывает на активацию нейронального ростка дифференцировки, т.е. происходит не только стимуляция образования новых глиальных элементов, но и нейронов. Период функциональной компенсации характеризовался постепенным восстановлением нейрональной плотности в ацетил-холин- и серотонинсинтезирующих ядрах, в которых через 6 мес. после удаления обонятельных луковиц уже не обнаруживалось ухудшения морфофункционального состояния нейронов. На рис. 8 представлена динамика восстановления плотности нейронов в структурах, синтезирующих серотонин, ацетилхолин и норадреналин.
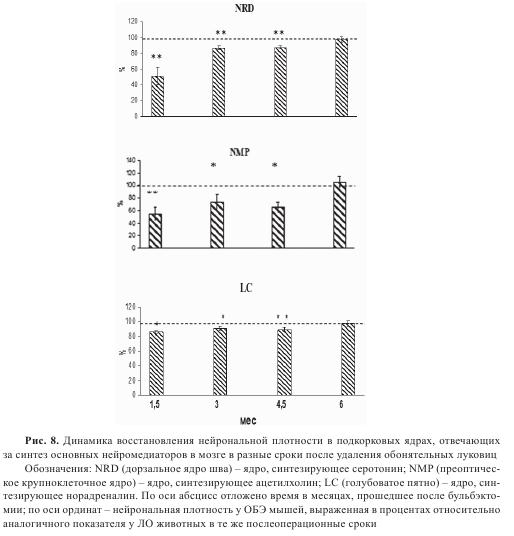
В мозге ОБЭ животных в этот период нормализовался уровень серотонина и увеличивалось число нейронов, позитивных к антителам против холинацетил-трансферазы, в базальных структурах переднего мозга. Восстановление серотонинергической функции должно способствовать дальнейшему процессу нейрональной дифференцировки прогениторных клеток через активацию синтеза мозгового нейротрофического фактора BDNF. Важно отметить, что обе эти системы через активацию 5-НТ2а или 5-НТ2с рецепторов, а также через мускариновые рецепторы, связанные с активацией фосфолипаз и киназ, способствуют активации α-секретазы, ответственной за неамилоидогенный путь метаболизма APP. Возможно, восстановление функционирования этих систем является одним из факторов, ответственных за снижение уровня Аβ, которое мы наблюдали на этапе компенсации у ОБЭ животных. Детальные исследования на трансгенных животных с усиленной продукцией APP в разные возрастные периоды показали активацию нейрогенеза у молодых животных и его снижение по мере накопления в их мозге Aβ. Однако, по данным других исследователей, Aβ из сенильных бляшек в основном стимулирует нейрогенез, но не глиогенез. Наши и литературные данные свидетельствуют, что активация нейрогенеза является одним из компенсаторных механизмов, используемых организмом, чтобы противодействовать развитию нейродегенеративного процесса на этапе стертой формы течения патологии. Для проверки данного предположения нами были выполнены опыты с трансплантацией фетальной ткани обонятельной луковицы, содержащей нейрональные прогениторы, в мозг ОБЭ животных через 7 мес. после бульбэктомии, т.е. в период, предшествующий манифестации патологии (рис. 9).

Эффективность трансплантации проявилась в улучшении памяти ОБЭ животных и нормализации морфо-функционального состояния нейронов височной коры и полей гиппокампа, в то время как ОБЭ животные, которым вводился физиологический раствор или ткань мозжечка, демонстрировали выраженную поведенческую и морфологическую патологию. Таким образом, эти данные свидетельствуют о компенсаторной роли активации нейрогенеза при развитии нейродегенерации и целесообразности его активации при БА.
Читайте также
Особенности сантехники из Италии: почему стоит переплатить

Отдых в Джемете — солнце, море, пляж

Джемете — это живописный район на побережье Черного моря, который
Школа профессионального макияжа: искусство создания идеального образа

-

Дорамы – популярный жанр телевизионного и кинематографического искусства, который нашел
Правильный выбор ткани — половина успеха вашего образа

Разработка брендбука: почему это важно

Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа
