Фармакологически вызванная хроническая гиперпролактинемия
Помимо наиболее широко используемой модели дегенерации катехоламинергических нейронов была разработана модель избирательной дегенерации только ДА-ергических нейронов при сохранении НА-ергических аксонов в аркуатном ядре у крыс. Это делает возможным оценить вклад в развитие гиперопролактинемии каждой из катехоламинергических компонент — ДА-ергической и НА-ергической. Дегенерация только ДА-ергических нейронов тубероинфундибулярной системы моделируется путем внутрижелудочкового введения 6-ГДА после предварительного системного введения десметилимипрамина (ДМИ) — протектора НА-ергических нейронов. ДМИ связывается с мембранным переносчиком НА на НА-ергических нейронах и таким образом ингибирует обратный захват НА, предотвращая захват 6-ГДА и дегенерацию нейронов. В отличие от предыдущей модели с введением только 6-ГДА, на этой модели не происходит компенсаторного восстановления уровня ПРЛ в крови (табл. 4). В результате концентрация ПРЛ в крови сохраняется в 2 раза повышенной как на 14-й, так и на 45-й день после введения 6-ГДА с ДМИ (табл. 4). В гипофизе наблюдаются отчасти схожие с предыдущей моделью изменения: содержание ПРЛ снижено на 14-й день на 65%, а на 45-й день на 30%, вес гипофиза до 45-го дня отстает на 20% от веса гипофиза контрольных особей, а содержание мРНК Д2-рецепторов уменьшается на 30%, причем только на 45-й день (см. табл. 4). Кроме того, через 48 часов после введения 6-ГДА и ДМИ, так же как и на модели введения только 6-ГДА снижается, митотическая активность лактотрофов, а через 96 и 144 часа повышается.
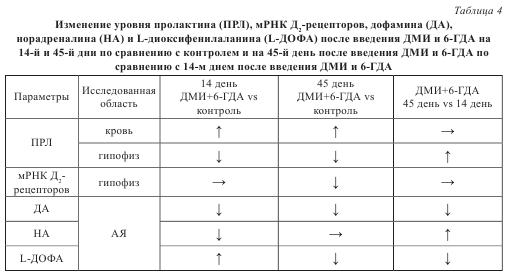
На данной модели изменения в уровне ПРЛ в крови коррелируют также с изменениями уровня ДА в гипоталамусе. В срединном возвышении и стебле гипофиза на 3-й день после введения 6-ГДА с ДМИ наблюдается снижение ДА на 50% без изменений содержания НА. В аркуатном ядре содержание ДА на 14-й день снижается на 75%, а на 45-й день — на 90%. При этом содержание НА незначительно (на 20%) снижается только на 14-й день и полностью восстанавливается к 45-му дню (см. табл. 4).
Для оценки секреторной активности нейронов аркуатного ядра также применяли метод проточной инкубации слайсов ткани на 14-й и 45-й дни после введения нейротоксина и в контроле (Дильмухаметова и др., 2012, 2014). При введения ДМИ с 6-ГДА на 14-й день НА выделялся спонтанно и стимулирование, но уровень выделявшегося НА был ниже, чем в контроле (см. табл. 5). При этом НА-ергические нейроны отвечают на калиевую-стимуляцию (см. табл. 5). Кроме того, суммарный уровень НА при введении ДМИ с 6-ГДА почти не отличается от контрольного (см. табл. 5). Эти результаты отчасти согласуются с данными других авторов, продемонстрировавших, что концентрация НА в срединном возвышении не изменялась через 24 часа после внутривенного введения ДМИ и 6-ГДА. Таким образом, наблюдается незначительное снижение НА-ергической иннервации с последующим частичным восстановлением.

В норме стимулированное выделение ДА нейронами аркуатного ядра в 3 раза выше, чем спонтанное. На 14-й день при введении ДМИ и 6-ГДА, так же как и при введении только 6-ГДА, было показано, что уровень спонтанного выделения ДА усиливался, а стимулированное выделение ДА отсутствовало, и в ткани уровень ДА был значительно снижен по сравнению с контролем (см. табл. 5). Также наблюдалось значительное снижение спонтанного и стимулированного выделения ДА по сравнению с контролем, но при этом ДА-ергическая система восстановила способность выделять ДА в ответ на калиевую стимуляцию, по сравнению с тем, что наблюдалось на 14-й день после введения токсина (см. табл. 5). Таким образом, на 45-й день наблюдается восстановление функциональной активности ДА-продуцирующих нейронов. Однако уровень ДА в ткани после инкубации на 45-й день после введения только 6-ГДА был выше, а после введения ДМИ с 6-ГДА — значительно ниже, чем в контроле. Это свидетельствует о том, что НА оказывает ингибирующее влияние на синтез ДА.
На 45-й день после введения ДМИ с 6-ГДА уровень спонтанного и стимулированного выделения L-ДОФА значительно ниже, чем в контроле (см. табл. 5). Кроме того, суммарное содержание L-ДОФА в ткани и среде после введения ДМИ с 6-ГДА меньше, чем после введения только 6-ГДА. Это свидетельствует о том, что уровень синтеза L-ДОФА ниже при сохранении НА-ергической иннервации аркуатного ядра, чем при ее отсутствии.
Биохимические изменения уровня ДА и НА в аркуатном ядре при введении ДМИ и 6-ГДА коррелируют с морфологическими изменениями в гипоталамусе после ведения нейротоксина. На модели введения ДМИ и 6-ГДА, так же как и на модели введения только 6-ГДА, наблюдалось снижение числа тирозингидроксилаза-иммунопозитивных нейронов в дорсомедиальной области аркуатного ядра, где локализованы в основном биферментные ДА-ергические нейроны, что объясняет значительное снижение синтеза ДА на обеих моделях как на 14-й день, так и на 45-й день (табл. 6). Однако в дорсомедиальной области оптическая плотность нейронов, коррелирующая с внутриклеточным содержанием тирозингидроксилазы, и площадь самих нейронов увеличивается при сохранении НА-ергического контроля (см. табл. 6). Это подтверждает предположение о протекторном влиянии НА по отношению к ДА-ергическим биферментным нейронам, которые локализованы в этой части аркуатного ядра.

При сохранении НА-ергической иннервации после введения ДМИ с 6-ГДА число тирозингидроксилаза-иммунопозитивных нейронов и содержание тирозингидроксилазы в нейронах в вентролатеральной области не изменилось (см. табл. 6). Приведенные данные свидетельствуют о том, что НА ингибирует синтез тирозингидроксилазы в моноферментных нейронах вентролатеральной области аркуатного ядра. Это хорошо согласуется с данными Дайкоку с соавторами, показавшими, что хирургическая деафферентация медиобазального гипоталамуса приводит к увеличению числа тирозингидроксилаза-иммунопозитивных нейронов в вентролатеральной области аркуатного ядра. Прямое доказательство НА-ергического ингибиторного контроля экспрессии тирозингидроксилазы в неДА-ергических нейронах было недавно получено нами при изучении вазопрессинергических нейронов супраоптического ядра, эксперсиирующих тирозингидроксилазу.
Из приведенных наших и литературных данных следует, что НА оказывает ингибирующее влияние на экспрессию тирозингидроксилазы в моноферментных нейронах вентролатеральной области аркуатного ядра и протекторное влияние на биферментные нейроны дорсомедиальной области аркуатного ядра.
Таким образом, при функциональной недостаточности ДА-ергических нейронов сохранение НА-ергической иннервации аркуатного ядра приводит к снижению экспрессии тирозингидроксилазы в неДА-ергических моноферментных нейронах аркуатного ядра и ингибированию синтеза ДА, что приводит к развитию хронической гиперпролактинемии.
Читайте также
Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа

Когда начинать готовиться к ЕГЭ и ОГЭ 2024: полезные рекомендации

Начало нового учебного года часто становится временем повышенной тревожности как
На чем можно долететь до Мальдив? Регулярный рейс или аренда частного самолета?

Путешествие на Мальдивы — это мечта многих туристов. Острова, утопающие
Зубной имплантат: преимущества выбора при протезировании

Зубной имплантат – это современная технология, предоставляющая возможность восстановить утраченный
Яйцо шоколадное Kinder сюрприз: волшебство, которое завоевало сердца детей и взрослых

Яйцо Kinder сюрприз, безусловно, является одним из наиболее популярных шоколадных
Суши и пицца: почему они так популярны в службе доставки

Службы доставки еды становятся всё популярнее среди людей, желающих насладиться
Пептидные препараты: сущность и области применения

Пептидные препараты стали одним из важнейших направлений в современной медицине