Дофаминергическая система среднего мозга и ее роль в регуляции моторного поведения
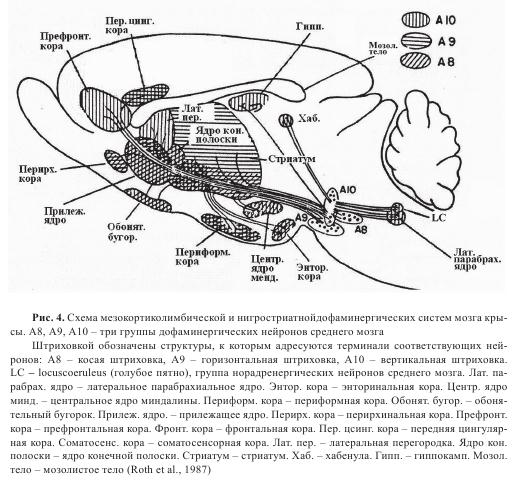
Известно, что ДА-нейроны среднего мозга объединены в три группы клеток: А8, А9 и А10. Группа А10 ДА-нейронов (рис. 4) локализована в вентромедиальной области покрышки среднего мозга (ВМО). Несколько латеральнее ДА-ергические нейроны компактной части черной субстанции формируют клеточную группу А9 (см. рис. 4). Нейронная группа А8 занимает более каудальную позицию в ретрорубральном поле среднего мозга (см. рис. 4). Терминали этих групп нейронов формируют проекции, которые образуют нигростриатную и мезокортиколимбическую ДА-ергические системы мозга. Нигростриатная система формируется в основном нейронами группы А9, тела которых расположены в компактной части черной субстанции. Терминали аксонов этих нейронов заканчиваются в области хвостатого ядра, или неостриатума. Мезокортиколимбическая ДА-ергическая система в большей своей части формируется телами нейронов А10. Аксоны мезокортиколимбической ДА-системы оканчиваются в вентральной части стриатума — прилежащем ядре, обонятельных бугорках, центральном и базолатеральном ядрах миндалевидного комплекса, в перегородке, гиппокампе, в префронтальной, передней цингулярной, энторинальной, периформной корковых областях и в глубоких слоях фронтальной коры.
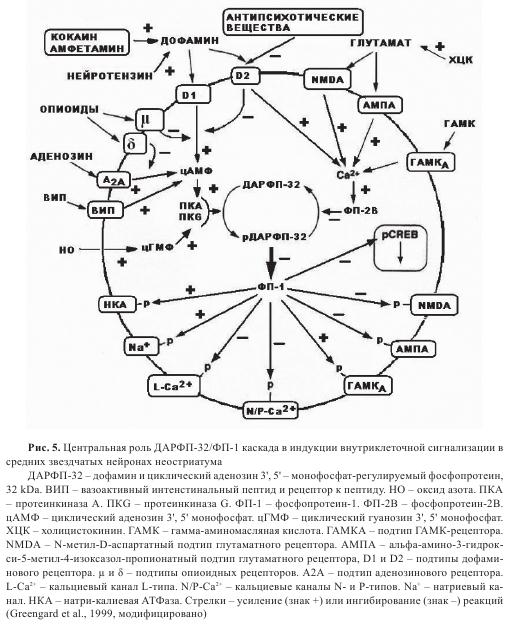
Как указано выше, нигростриатная ДА-система контролирует активность ГАМК и глутаматергических синапсов неостриатума, которые, в свою очередь, активируют таламокортикальные сети и регулируют моторное поведение животных и человека. Подробно, индукция внутриклеточных сигналов ДА-рецепторами и контроль активности ГАМК и глутаматергических синапсов средних звездчатых нейронов неостриатума описаны в работе П. Грингарда и соавторами. Как следует из предложенной схемы (рис. 5), ДАРФП-32/ФП-1 (ДА регулируемый фосфопротеин 32 килодальтон/фосфопротеин-1) интегрирует информацию от ряда нейротрансмиттеров и вызывает координированные реакции, включающие работу некоторых выходных физиологических эффекторов. Фосфорилирование ДАРФП-32 через протеинкиназу А (ПКА) регулируется действием разных нейротрансмиттеров. ДА действует через D1-рецептор, аденозин действует через аденозиновый А2А-рецептор, вазоактивный интенстинальный пептид (ВИП) — через соответствующий ВИП-рецептор. Протеинкиназа G (ПKG) активирует ответ оксида азота (НО) также фосфорилированием ДАРФП-32. В звездчатых клетках среднего размера, которые экспрессируют D1- и D2-типа ДА-рецепторов, активация D2-pцепторов вызывает уменьшение уровня цАМФ. Опиоиды, действующие или через μ- или через δ-рецепторы, уменьшают уровень цАМФ в стимулируемых D1-агонистами клетках или аденозиновыми А2А-агонистами соответственно. Фосфорилированный ДАРФП-32 дефосфорилизируется ФП-2В и Са2+-калмодулинзависимой протеинфосфотазой. Фосфопротеин-2В (ФП-2В) активируется рецепторами нескольких нейротрансмитеров, в основном опосредованных Са2+-током NМDА-рецептора. Действие глутамата через АМРА-рецептор также стимулирует дефосфорилирование ДАРФП-32 через ФП-2В, который вовлекается деполяризацией нейрона и Са2+-током. Активация D2-рецепторов также ведет к увеличению уровня Са2+ через неидентифицированный механизм и увеличивает активность ФП-2В. Наоборот, действие ГАМК через ГАМКА-рецептор стимулирует ДАРФП-32 фосфорилирование гиперполяризацией нейрона, уменьшением Са2+-тока и инактивацией ФП-2В. Нейротензин усиливает выделение ДА и увеличивает фосфорилирование ДАРФП-32. Холицистокинин (ХЦК) усиливает глутаматергическую передачу и уменьшает фосфорилирование ДАРФП-32. Психомоторная стимуляция кокаином и амфетамином увеличивает фосфорилирование ДАРФП-32 усилением ДА-передачи. Все антипсихотические вещества достигают определенного клинического эффекта через антагонизм D2-peцепторов, ведущий к увеличению фосфорилирования ДАРФП-32. Фосфо-ДАРФП-32 ингибируется активностью ФП-1, действуя в синергической манере с разными протеинкиназами (в основном с ПКА и ПКС; на рис. 4 не показано), увеличивая уровень фосфорилирования различных эффекторных белков. В результате увеличение фосфорилирования ассоциируется с увеличением активности NMDA и АМПА рецепторов глутамата, L-, N-, и P-типа Са2+-каналов, и с уменьшением активности ГАМКА-рецепторов, Na+-каналов, и Na+/K+-АТФазы. ФП-1 фосфорилирует CREB, который является ранним геном, промотором экспрессии поздних генов. Таким образом, трансдукционный сигнал в конечном счете модифицирует экспрессию генов, которые консолидируют модификации, вызванные модуляторными реакциями.
Предполагается, что ГАМК и глутаматовые рецепторы взаимодействуют через единую интегрирующую систему, взаимодействуя через внутриклеточные метаботропные реакции, т.е. через трансдукционный сигнал, что экспериментально показано во многих исследованиях. Так, например, при ПТЗ-индуцируемом киндлинге редукция ГАМК-ергической функции блокируется антагонистом NMDA-рецепторов МК-801. NMDA-рецептор вовлекается в процесс формирования киндлинга, индуцируемого FG 7142, обратного агониста БДЗ-рецептора. Киндлинг, продуцируемый высокочастотной стимуляцией гиппокампа, индуцирует повышение активности NMDA-рецептора, также как модификацию ГАМКА-рецептора. В то же время, было обнаружено, что NMDA-индуцируемая долговременная потенциация может контролироваться регуляторными системами ГАМКА-рецепторного комплекса, ингибирование агонистами БДЗ-рецепторов, a также облегчение антагонистами тех же рецепторов. И наоборот, NMDA-антагонист CPP может модулировать долговременную модификацию (толерантность) БДЗ-рецептора. Возможная схема взаимодействия и интеграции ГАМКА и глутаматергических рецепторов на уровне внутриклеточной сигнализации через трансдукционный сигнал приведена на рис. 6.
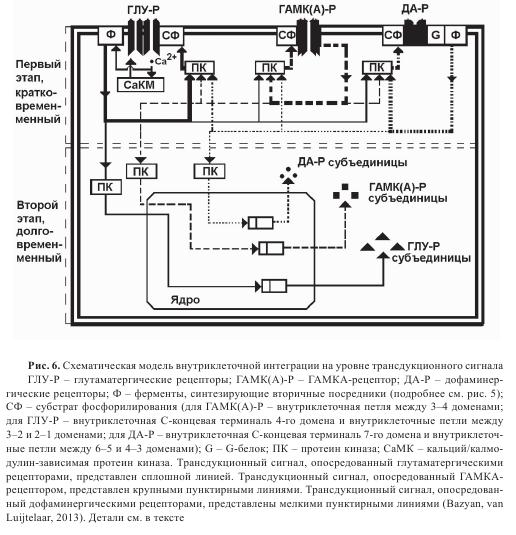
Возможно, что БДЗ-рецептор — один из сайтов ГАМКА супрамолекулярного рецепторного комплекса, запускает внутриклеточные метаболические реакции, который и является объектом долговременной модификации. Любой долговременно сохраняющийся процесс поддерживается модификацией экспрессии генов. БДЗ-рецептор, действуя через систему вторичных посредников и протеин киназу С, может индуцировать медленные метаболические реакции внутри клетки и модифицировать экспрессию генов в различных структурах мозга крыс. В частности, этот процесс в стриатуме приведен на рис. 5. Известно, что ГАМКА-рецепторы регулируются фосфорилированием опосредованном через протеин киназу С и G. Когда в нейронах активность протеинкиназных реакций усиливается, то хлорный ток, текущий через ГАМКА-рецептор, редуцируется.
Изменение уровня внутриклеточного фосфорилирования протеинкиназой может модифицировать глутаматергические рецепторы, повышая их ответ на эндогенные возбуждающие аминокислоты. Метаботропная регуляция глутаматергического синапса идентична описанным процессам, как в нейронах гиппокампа, так и в нейроных коры и стриатума. Мы добавили петлю обратной связи для метаботропной регуляции NMDA. Мы предполагаем, что механизм обратной связи метаботропной регуляции или ауторегуляция рецепторов через модуляторные реакции является необходимым условием для поддержания процессов долговременной потенциации или долговременной депрессии. Регуляция АМРА и ауторегуляция NMDA-рецепторов в гиппокампе были исследованы экспериментально.
Хорошо известно, что ДА-система мозга является системой поощрения и подкрепления, которая, как предполагается, впрямую вовлекается в процессы обучения и памяти. Кроме того, ДА-система вовлекается в формирование различных эпилептоформных состояний. Полученные нами результаты позволили нам предположить, что ДА-рецепторы и ГАМКА-рецепторы мозга взаимодействуют и модифицируют друг друга, интегрируясь и формируя зависимое от воздействий (обучение, судороги и так далее) эмоциональное состояние. Мы также предполагаем, что это взаимодействие и интеграция завершается внутриклеточной метаботропной интеграцией глутамат ГАМКА ДА-рецепторов.
Функциональное взаимодействие глутаматных и ДА-рецепторов стриатума были описаны в нескольких работах, в которых было обнаружено реципрокное или антагонистическое взаимодействие. Анализ большого количества результатов позволили Р. Коттеру предложить концепцию постсинаптической интеграции глутаматергических и ДА-сигналов в стриатуме на основании взаимодействия внутриклеточных вторичных посредников. Было показано, что индукция долговременной депрессии низкочастотной стимуляцией нейронов поля СА1 гиппокампа находится под влиянием обоих и NMDA- и ГАМК-рецепторов и оба D1- и D2 ДА-рецептора вовлекаются в модуляцию долговременной депрессии; при этом активация D1 ДА-рецептора усиливает долговременную депрессию, а активация D2 ДА-рецептора ингибирует долговременную депрессию. Ингибирование долговременной депрессии блокируется пикротоксином, но этот эффект приостанавливается SKF-38393-агонистом D1-рецепторов. Наши результаты также указывают на синергичное взаимодействие ГАМКА- и ДА-рецепторов при воспроизведении памяти.
Метаботропная интеграция глутамат ГАМК- и ДА-рецепторов схематически показана на рис. 6. ДА-рецепторы могут подвергаться аутомодификации петлей метадотропной обратной связи, когда модифицируют активность глутамат и ГАМК-рецепторов сигналами внутриклеточного фосфорилирования. С помощью той же самой реакции внутриклеточного фосфорилирования глутамат и ГАМК-рецепторы могут контролировать эффективности ДА-рецепторов. На втором этапе модификация, сформированная первым этапом, консолидируется модификацией экспрессии соответствующих генов.
Способность ДА-рецепторов подвергаться аутомодификации была продемонстрирована как на уровне связывания радиоактивных лигандов, так и на уровне экспрессии генов в различных структурах мозга и при различных экспериментальных процедурах.
Схема внутриклеточной интеграции на уровне трансдукционного сигнала для звездчатых нейронов неостриатума подробно описана. Обе схемы и наша, и Грингарда, в основном похожи, но есть несколько принципиальных отличий.
Схема Грингарда описывает только первый этап реакции, то есть в основном процессы фосфорилирования и дефосфорилирования. В то время как наша схема описывает двухэтапный процесс. Первый этап — кратковременный (минуты), процессы фосфорилирования и дефосфорилирования. Второй этап — долговременный (часы сутки месяцы), основанный на процессах модификации экспрессии генов.
Самое основное различие заключается в том, что наша схема описывает не только взаимомодификацию активности рецепторов, но и их аутомодификацию с помощью механизмов обратной связи. В случае с ГАМКА-рецептором аутомодификация с помощью обратной связи осуществляется через механизм аллостерической пластичности ГАМКА-рецептора.
Читайте также
Услуги гидроизоляции — внешняя или внутренняя?

Трудовые споры: как добиться справедливости от недобросовестного работодателя

Трудовые отношения — это тонкая материя, полная нюансов и правовых
Как отличить брендовые очки от подделки

Брендовые солнцезащитные очки — это не только модный аксессуар, но
Дизайн встроенной кухни: как оптимизировать пространство

Несмотря на большое разнообразие готовой (типовой) мебели, мебель на заказ
Михаил Владимирович Мишустин: отличный управленец и экономист

Михаил Владимирович Мишустин — выдающийся российский государственный и политический деятель,
Самые популярные рецепты пиццы: идеальное сочетание ингредиентов для настоящего гурмана

Пицца – это одно из наиболее популярных блюд в мире,
-

Лето – это время, когда дети, закончив учебный год, уходят
Как получить гражданство Бельгии и что оно дает?

Бельгия, расположенная в сердце Европейского союза, по праву считается одним
Осетинские пироги: вкусное и популярное блюдо с Кавказа

Когда начинать готовиться к ЕГЭ и ОГЭ 2024: полезные рекомендации

Начало нового учебного года часто становится временем повышенной тревожности как
На чем можно долететь до Мальдив? Регулярный рейс или аренда частного самолета?

Путешествие на Мальдивы — это мечта многих туристов. Острова, утопающие
Зубной имплантат: преимущества выбора при протезировании

Зубной имплантат – это современная технология, предоставляющая возможность восстановить утраченный
Яйцо шоколадное Kinder сюрприз: волшебство, которое завоевало сердца детей и взрослых

Яйцо Kinder сюрприз, безусловно, является одним из наиболее популярных шоколадных
Суши и пицца: почему они так популярны в службе доставки

Службы доставки еды становятся всё популярнее среди людей, желающих насладиться
Пептидные препараты: сущность и области применения

Пептидные препараты стали одним из важнейших направлений в современной медицине